Critical appraisal on the Helminth-Tuberculosis interaction
The Critical thinking bit....
Ezenwa
et al, 2015
OPPOSITE
EFFECTS OF ANTIHELMINTIC TREATMENT ON MICROBIAL INFECTION AT INDIVIDUAL VERSUS
POPULATION SCALES

Ezenwa
and Jolles (2015) thought deworming would lift the immune suppression by
helminth-infection resulting in BTB clearance in wild buffalos but that
wasn’t the case! Well….lets find out what really happened!
|
There is an increasing observation of the interaction between helminth and microbial infection which has led to the suggestion of integrating anthelmintic treatment and microbial disease control to improve disease outcome. This study aimed to investigate what effects such interventions would have on the target-microbial disease pattern at population level. Wild buffalo naturally infected with gastro-intestinal helminth were captured, treated and control group left untreated. It was observed that treated animals were less likely to be infected by BTB and shed fewer eggs than control group but there was no change in probability of BTB infection observed. However, treatment reduced mortality with higher IFN-y production which is essential in limiting BTB. In addition, anthelmint treatment is shown to have no impact on infection risk but rather continued spread of disease by the surviving BTB positive anthelmintic treated individuals thus a negative population level impact.
Treated buffalos survived longer than untreated
buffalos.

Ezwena et al (2015)
|
Noteworthy, adequate sample size of 216 buffalos was
used ensuring that all the buffalos were BTB infection free before exposer to
BTB and had the same likelihood of infection with helmints. Also, the
incomplete and inconsistent worm clearance by the anthelmintic treatment in
this study and the observation study design mimics the outcome of large scale
human deworming programs. However, the low number of eggs shed in treated group
than in the control group was subject to the length of the capture period thus
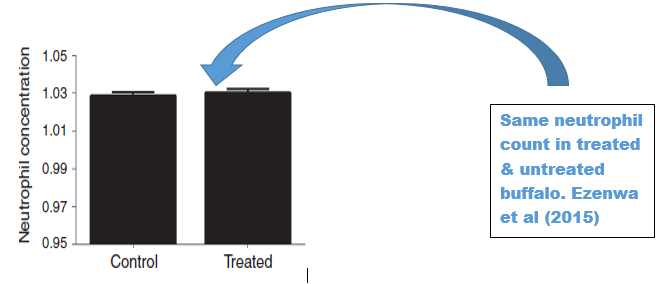
not a reliable criteria. The study found no difference in the neutrophil counts between
treated and the control group in the buffalo model yet in humans differences in
neutrophil counts are known to occur where low neutrophil counts are associated
with higher risk of BTB infection and impaired restriction of BTB growth
indicating difference in responses in the buffalo animal model and human which
is a challenge in study models (Kanistanon et al 2008).
Ezwena et al (2015)
POTIAN et al, 2011
PRE-EXISTING HELMINTH INFECTION
INDUCES INHIBITION OF INNATE PULMONARY ANTI-PULMONARY ANTI-TUBERCULOSIS DEFENSE
BY ENGAGING THE IL-4 RECEPTOR PATHWAY
 Lately,
a lot of literature is building up on the interaction between Helminth and MTB infection, therefore, Potian
et al decided to investigate whether the immunological environment created by a
preexisting helminth infection has an impact on the host protective response
against MTB infection. BALB/c
mice were subcutaneously infected with L3 stage Nb larvae, 5 days later, these mice were
aerosolized with Erdmann strain MTB
and examined for outcomes such as MTB burden, T cell and Granuloma responses etc. It was observed that L3
larvae induced a strong TH2 response
in the lungs that caused a transient impairment of resistance to MTB infection
around week 4 although the TH2 response did not affect the TB specific TH1 response since IL-4 and IFN-y populations did not
differ in the co-infected and control groups.
Lately,
a lot of literature is building up on the interaction between Helminth and MTB infection, therefore, Potian
et al decided to investigate whether the immunological environment created by a
preexisting helminth infection has an impact on the host protective response
against MTB infection. BALB/c
mice were subcutaneously infected with L3 stage Nb larvae, 5 days later, these mice were
aerosolized with Erdmann strain MTB
and examined for outcomes such as MTB burden, T cell and Granuloma responses etc. It was observed that L3
larvae induced a strong TH2 response
in the lungs that caused a transient impairment of resistance to MTB infection
around week 4 although the TH2 response did not affect the TB specific TH1 response since IL-4 and IFN-y populations did not
differ in the co-infected and control groups.
Potian et al (2011)

| Potian et al (2011) |
However,
reinfection with helminth is shown to increase MTB burden by two fold with
larger granuloma formation, high macrophage-alternative activation and
mortality in 5 weeks in mice. The investigations in this study were carried out
as independent experiments in triplicates or duplicates and clearly indicate
that pre-existing helminth infection impairs host ability to contain MTB
infection by alternative activation of macrophages.
However,
use of 500 L3 larvae for subcutaneous injection of the mice way-above the
normal infection dose in natural helminth infection. Use of such very high dose
of infection may have modified the responses observed since very high or very
low doses of Invitro helminth infection studies
have been seen to elicit different immune responses. To determine numbers of
AAMs and CAMs in the co-infected and control-group, FIZZ-1 was used as a marker for AAMs yet ARG-1 is a more specific
marker for AAM. However, the article admits difficulty of flow cytometry
staining for ARG-1 and thus specificity of ARG-1 marker was catered for by
first gating the cells based on their CD11b and c expression. This study also proves that alternative
macrophage activation occurs with or without IL4R engagement but did not enumerate mechanisms
that lead to activation of AAMs without IL4R engagement. Noteworthy, the mechanism of how AAM supports
MTB growth was not proven but rather based on a conjurement that AAMs have a
reduced nitrosative stress. A high number of AAMs observed in week 4 during the
transient impairment of protection from MTB and later high CAM numbers in week
7 that restored protection was observed, however, whether this observation was
due to conversion of the AAMs to CAMs or due to the CAMs being independently
activated after apoptosis of the AAMs remains unanswered.
In conclusion, these studies by Ezenwa et al (2015) and Potian et al (2011), demonstrate that helminth infection elicits TH2 response and increases alternative-activation of macrophages resulting in less controlled MTB growth, increased infection severity and mortality. However, on anthelminth treatment, AAM induction is reduced while CAM induction is increased thus MTB infection burden is controlled and reduced. This reduces individual’s mortality but the incomplete helminth clearance by the anthelminth treatment still results in MTB transmission in the population.
References
No comments:
Post a Comment